Restoring the Kalligrammatids: The Not-Butterflies of the Mesozoic

Recently
I’ve been working on a set of illustrations featuring the kalligrammatids, a
family of Mesozoic lacewings which are most well known for their butterfly-like
appearance and wing eyespots, and while I’ve been doing it I’ve realised that
there are no good references or guides to their life appearance available to
palaeoartists and people just interested in these insects. So, while this is by
no means a comprehensive guide, here are a few quick notes on what I’ve learnt
about the kalligrammatids and how to restore them. I’d like to start out by
saying that I am not an invertebrate specialist; as some of you will know, and
many of you will be able to guess from the name of my blog, I am a
palaeobotanist, so my specialist knowledge is on plants, not insects, so if any
of you more adept in the ways of the bug notice anything glaringly wrong here
please let me know!
It is a common trend within palaeoart
that certain groups get WAY more attention than others (I’m looking at you, non-avian
dinosaurs), and any group which is even the tiniest bit obscure often has no
easily accessible information about it. Kalligrammatidae is one of these groups.
Yet despite this they are reasonably common elements within palaeoart (albeit
often merely as background elements in a scene featuring something larger and
toothier). This is unsurprising, as they are some of the showiest insects of
the Mesozoic, and a number of well-known studies with a lot of news coverage have been released in the last ten years or so (Labandeira et al., 2016;
Yang et al., 2014). However, despite their relatively common inclusion
within Mesozoic scenes, very few people have explored the diversity of
kalligrammatids which were present. An unfortunate trope when restoring
kalligrammatids is to make them almost butterfly-clones; this is not that
surprising, as they have been ‘marketed’ as being very butterfly-like, and
indeed they are (I was actually a little surprised by how superficially similar
some taxa are to modern lepidopterans)! However, this similarity is only very
superficial, and closer examinations of their fossils and other neuropterans (the
net-winged insects, the group to which the kalligrammatids belonged) reveals
that their anatomy was rather different from butterflies.
So, what are the kalligrammatids?
Well,
first off, let me introduce you to the kalligrammatids: the kalligrammatids are
an extinct family of winged insects within the order Neuroptera,
a clade which in the modern world includes things like lacewings, mantidflies (Mantispidae)
and antlions (Myrmeleontidae). They
originated in the late-Early Jurassic (~175-180 Ma) (Ansorge and Markarkin,
2020) and went extinct in the mid-Early Cretaceous (~124 Ma) (Bechly and Makarkin,
2016; Yang et al., 2014; Zhong et al., 2021). Almost all species
originate from Eurasia, although three species are also known from the Crato
Formation of northern Brazil, suggesting their distribution was larger than is
currently known (Machado et al., 2021). Several small neuropterans have
been found in the Myanmar amber, however their taxonomy is uncertain; these
insects were initially considered to form their own clade, outside of
Kalligrammatidae, but within the same superfamily (Psychopsoidea, of which the
only extant family is Psychopsidae, the silky lacewings)
(Lu et al., 2016), however, a subsequent study found that these, and
several new genera, formed a small subfamily (Cretanallachiinae) within
Kalligrammatidae (Liu et al., 2018). If these small neuropterans do turn
out to be kalligrammatids, this would extend the known geological and chronological
range of the family, however, because of their taxonomic uncertainty (and slightly
different appearance) I will not be discussing them other than to point out the
following:
·
They
are much smaller than other known kalligrammatids; almost 1/10th the
size of even the smallest kalligrammatids I’ve illustrated here.
·
The
males have bipectinate (frilly, comb-like) antennae (which aren’t known from
other kalligrammatids; Yang et al., 2014).
·
They
have elongated proboscis-like mouthparts so were nectar feeders.
·
They
have two pairs of palps (feeding appendages), maxillary and labial, unlike
other kalligrammatids which appear to only have one pair (Labandeira et al.,
2016 supp. Info.).
·
And
some species had eyespots on their wings (e.g. Burmogramma liui),
although most have clear wings (Liu et al., 2018).
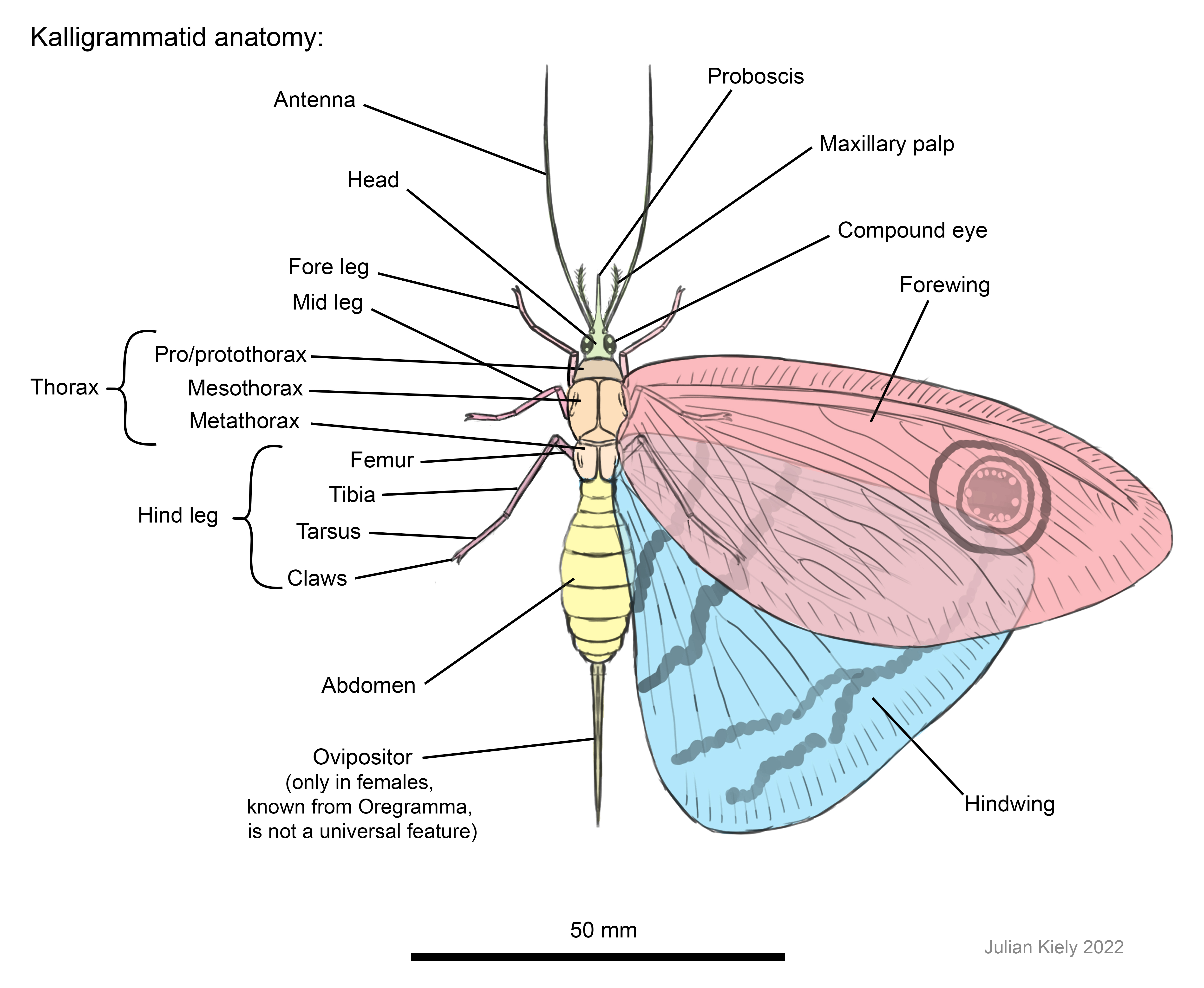
Kalligrammatid anatomy
When
doing my recent illustrations, I started out by producing an anatomical diagram
of one species, so that I could get to grips with their anatomy and life appearance
(Fig. 2). This was interesting to do, as I’m not aware of this having been done
before, so my restoration is based mainly on the fossils we have (which often
preserve the wings in a lot of detail, but with disarticulated bodies), with
other details supplemented by looking at modern neuropterans. Some of the main
points which I found important when restoring kalligrammatids are as follows:
·
Their
thorax (the middle section of their body) is split into three sections (the pro
or protothorax, the mesothorax, and the metathorax) and the forewings attach to
the mesothorax, and hindwings to the metathorax.
·
The two
pairs of wings often overlap and the forewings overlap the hindwings dorsally (when
viewed from above the forewings are on-top).
·
The
bases of their wings are cordate where they attach to the body, meaning that
they’re heart-shaped, with two bulges, in-front of and behind the point of attachment
– forgive me for using a botanical term (‘cordate’), there may be a formal term
for this in insect wings, but I don’t know it.
·
Their eyes
were often large, sometimes almost meeting in the middle of the head.
·
Their
wings were covered in microscopic scales, which would have given the wings
colour; however…
·
Their wings
were likely most often translucent, like most modern neuropterans, as most
species do not seem to have a thick covering of scales; however…
·
Some species
DID have a thick covering of scales, which in the fossils completely, or
near-completely obscure the venation, and these taxa likely had opaque wings (e.g.
Abrigramma calophleba; Sophogramma lii, Ithigramma spp.
and Meioneurites spectabilis).
·
Their
antennae are filiform (thread-like and segmented) and often long, though not as
long as the forewings, and short in some genera (e.g. Ithigramma).
·
Their
bodies likely had a sparce covering of hairs, although these may have been
localised and some diversity in the distribution, density and length of these
hairs would be expected (I have not observed long hairs in any fossils, so
their bodies would not have been fluffy like moths; you can look at modern
neuropterans for inspiration).
·
Their
legs may also have had a sparce covering of hairs.
·
They
probably didn’t have long tibial spurs, although short ones may have been
present in some species (e.g. they’re known from the fore legs of Meioneurites
spectabilis). As a side note, this was something I only noticed while
writing this, after having finished all the diagrams, so I had to go and erase them
from all my drawings (apologies if I missed some).
·
Long, curved
ovipositors are known from at least one species (Oregramma illecebrosa),
but they seem to be absent in most taxa, even when the specimens are presumably
female and well preserved. This means that kalligrammatids should not be
ubiquitously restored with long ovipositors.
·
The length
of their abdomens varies, but none extend beyond the length of the hindwings,
and the abdomens of males were likely very short (see Oregramma sp.;
Yang et al., 2014).
·
Most kalligrammatids
had a proboscis (siphonate mouthparts), with mandibulate mouthparts being
restricted to basal clades (Sophogrammatinae).
The diversity of nectar-feeding
kalligrammatids
The
thing which started my recent obsession with kalligrammatids was when I began
doing an illustration showing the diversity of species which would have fed on
nectar and been plant pollinators (Fig. 3). All of the species I have restored
are known to have had a proboscis, or siphonate mouthparts (Yang et al.,
2014) with the exception of Markarkinia, which is only known from wing
fragments. I included two species from this genus to show that some
kalligrammatids could get very large; the largest are estimated to have had a
wingspan of ~30 cm across, with individual wings being 16 cm long (Machado et
al., 2020), however here I have been a little more conservative, giving
them only ~25 cm wingspans. This is something which I was consistently surprised
by when restoring these kalligrammatids: exactly how large most of them
where!!! Of the species which I looked at in detail, the smallest ones still had
wingspans of about 9 cm, the size of the UK’s largest butterfly, the
swallowtail, and most had wingspans in the range of 12 to 19 cm across, much
larger than any modern neuropterans, which have wingspans maxing out around 10 cm
across. These large sizes are quite alarming for insects and is something worth
considering when restoring kalligrammatids. It is also worth thinking about
when pairing kalligrammatids with plants; in fact, it may help explain why some
bennettitalean ‘floral organs’ were so large (e.g. Weltrichia sol had a ‘floral
organ’ up to 20 cm across; Popa, 2019).
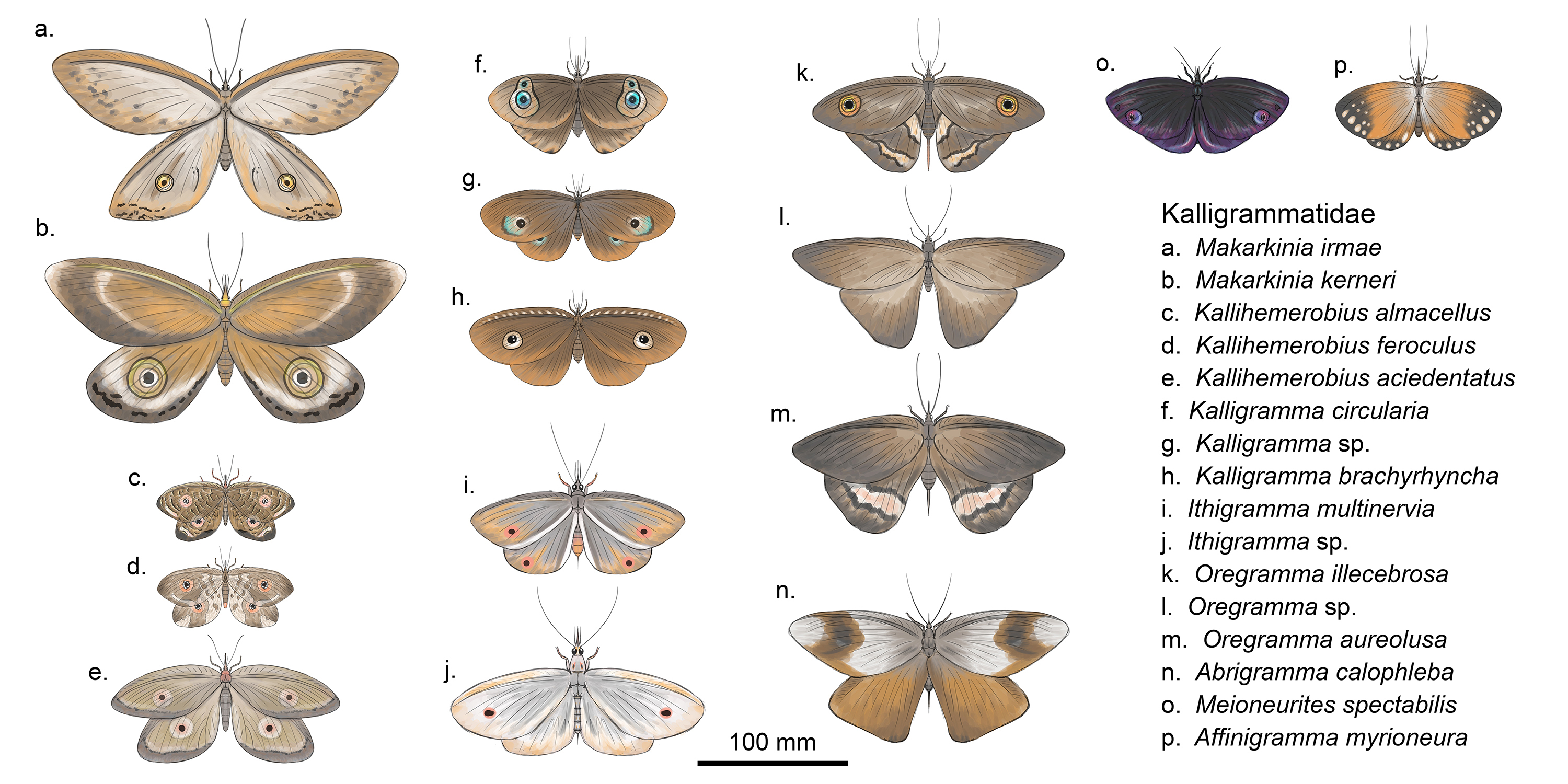
Something else I found fascinating
when restoring the kalligrammatids was just how morphologically diverse they
actually were. I am so used to seeing the one or two different restorations
being used over and over again that I was surprised at how much their appearance
varied from species to species. The most obviously variable thing about them
was their wing shapes (see Fig. 3); as a general rule, the forewings are always
longer than the hindwings, and often narrower, with a higher aspect ratio, and
the hindwings were much broader and often rounded or almost triangular-shaped. However,
some species (e.g. Meioneurites spectabilis and Affinigramma
myrioneura) had forewings which were incredibly broad and almost completely
hid the smaller hindwings, creating an almost skirt-like silhouette when the
wings were spread out. Still other species, such as those within Kallihemerobius
possessed hind and forewings which were almost identical in size and shape. In Oregramma
and Abrigramma the forewings are very long and narrow with pointed
tips, and their hindwings are triangular-shaped and large.
One of the most unexpectedly
variable features within the kalligrammatids was the length and size of their
prothorax (also called a protothorax; Fig. 2). The prothorax is the first
segment of their thorax, and in some modern neuropterans serves almost as a neck,
which can be moved up and down relative to the rest of the body (the forelegs
also attach to this body section). In all kalligrammatids the top of the prothorax
appears to be almost shield-like, with a smooth curved surface (actually, now
that I think about it, it’s more saddle-like) which extends down its sides. In
most species it is pretty innocuous, being similar to that seen in Oregramma
illecebrosa in Fig. 2; however, in some species it is incredibly long and
robust, causing the head to project out in front of the wings! This is most
obvious in Ithigramma sp. (Yang et al., 2014), although it can
also be observed to a lesser extent in Oregramma sp. and Abrigramm
calophleba. Meioneurites spectabilis also has a longer, but also
narrower, prothorax. Variability in the length of the prothorax is a feature
also commonly seen in modern neuropterans, with extreme examples being seen in
the mantidflies and snakeflies,
so it’s interesting that the prothorax of kalligrammatids is also very
variable.
The shape and size of kalligrammatid
abdomens does vary somewhat, although most are short and broad. Some genera did
possess longer, narrower abdomens, particularly Kallihemerobius (which
had long abdomens which were broadest near their base and narrowed gradually towards
their end) and Meioneurites (which had an abdomen which almost extended
beyond the back-edge of the wings and was an elongate oval shape with a blunt
end; Engel, 2005). Some kalligrammatids possessed long, curved ovipositors, as
seen in Oregramma illecebrosa, but the distribution of these within the
group is not known, and it likely was not a ubiquitous feature, as multiple
taxa with well preserved abdomens show no sign of an ovipositor (although the
presence/absence of ovipositors is also confused by the fact that male
specimens are difficult to identify as male unless the preservation is superb)
(Engel, 2005; Yang et al., 2014). As a general rule I would say, if in
doubt, don’t include an ovipositor when restoring kalligrammatids.
The head shape of most
kalligrammatids is pretty consistent, although details such as the length of
the proboscis and palps, antenna length and overall head size do vary somewhat.
These variations can be seen in Fig. 3. (although I appreciate at this scale it’s
a little difficult to see). As mentioned earlier, several taxa of
kalligrammatids (e.g. Sophogramma lii) did have mandibulate
mouthparts, however I have not looked at these taxa in detail, so cannot talk
about their appearance.
Leg length was also incredibly
variable within the kalligrammatids, although this cannot be discussed in much
detail as few specimens preserve legs in any great detail. Most taxa with preserved
legs seem to have reasonably short ones (e.g. Ithigramma and Abrigramma)
which ended in two curved claws and (seemingly) lacked large tibial spurs (Yang
et al., 2014). The legs of Meioneurites spectabilis, while
also short, were very robust, and possessed short tibial spurs on at least the
first two pairs of legs; it also possessed large claws at the end of its short robust
tarsus. Another species worth discussing is Kallihemerobius almacellus;
while the legs of this species are not very well preserved, they indicate that
this species had very long legs (Yang et al., 2014).
Perhaps the most beautiful feature
of at least the fossils of kalligrammatids is that most preserve at least some
patterning! These patterns most commonly (and most famously) take the form of
eyespots, which are present within almost all kalligrammatids. Because of their
wide distribution, it is unsurprising that the forms of these eyespots vary
quite significantly between the different genera, and even between species of
the same genera. Five different forms of eyespots (named Type 1 - 5) were
described by Labandeira et al. (2016) and these range from simple black
spots, as seen in Ithigramma and some species of Kallihemerobius,
all the way to complex ‘eyes’ which incorporate eyeshine-spots and multiple
rings, like those in Oregramma illecebrosa, Makarkinia, Kalligramma,
and some species of Kallihemerobius. One of the most fascinating wings I
came across was that of Kalligramma brachrhyncha. When restoring
this species I noticed that the holotype preserves a series of fain pale slanting-triangles
along the anterior edge of the wing, which in the fossil appear pretty
innocuous; however, when adding these pale patterns to my restoration (Fig. 3h,
5) I was surprised when I was greeted by a toothy grin! While a closer
examination of the fossil would be needed to confirm that this pattern is
genuine, this could represent a remarkable level of mimesis, with the wings mimicking
the head of a small predator (small dinosaur anyone?) when folded closed.
Something else which I found interesting when restoring multiple genera is that
similar genera often share similar types or distributions of eyespots. This is
unsurprising, but still interesting to see such consistent patterns within
fossil genera; this consistency is useful when restoring incomplete species
from the same genera. For instance, the fore and hindwings of Kallihemerobius
aciedentatus are known and they seem to indicate that both fore and
hindwings had eyespots (although Labandeira et al., 2016 interpreted the
fossil as having two spots on the forewing only); in the other two species of Kallihemerobius
which I restored (K. almacellus and K. feroculus) only either the
fore or hindwings are known from each species, however both possess complex
eyespots so it is reasonable to assume that eyespots would have been present on
both fore and hindwings, like in Kallihemerobius aciedentatus. In Ithigramma
species, the eyespots are simple black spots and appear to be present on both
the fore and hindwings of both species (although they are obscured by the
forewings in Fig. 3j). The eyespots in Kalligramma species are quite
variable, but consistently have a central black spot with smaller
eyeshine-spots which are surrounded by a pale region enclosed within a dark
ring. The eyespots of Kalligramma circularia are difficult to interpret
from the fossil, but this species either has two eyespots per wing (as I have
shown here) or they had a single eyespot on the forewing, and a smaller one on
the hindwing. Both Makarkinia species which I have shown are known from
the hindwings, and both possess large eyespots with a simple black centre
encircled by three dark rings of varying thickness. Finally, in Oregramma
only one species seems to preserve an eyespot, however the hindwings of both Oregramma
illecebrosa and Oregramma aureolusa seem to have light and dark
barring. As previously mentioned, the wings of both Abrigramma and Meioneurites
were very densely covered in scales, so it is likely that in life the wings of these
species were opaque, possibly with very rich colouration. Simple dark eyespots
are apparently present in Abrigramma calophleba (Labandeira et al.,
2016), however I could not identify them or their position in the fossil. Affinigramma
was the only genus I restored for which I could find no evidence of eyespots.
Kalligrammatid flight (probably
wasn’t butterfly-like)
As
previously mentioned, the kalligrammatids were neuropterids, so the following
section is mainly based on observations of modern neuropterid flight, and then
applying that to the kalligrammatids. When thinking about kalligrammatid flight
I looked mainly at videos on youtube showing the take-off and flight of a
variety of modern neuropterids. Of particular use were videos by the users NickSchrader and Ant Lab (video one and two).
I’ve outlined my conclusions below (and in a rather janky animation I made; Fig.
4), but it is also worth pointing out that this is my own interpretation, and I’m
not a biomechanist by any means:
·
When
taking flight the kalligrammatids would start by unfolding their wings and
spreading them out.
·
They would
then raise their wings up while also tilting up their prothorax, lifting their front
pair of legs off the ground (this is something which all modern neuropterans apparently
do, as it moves the legs out of the way of the wings).
·
The
kalligrammatids would then flap down, moving their forewings down, followed by
their hindwings a moment later.
·
This downstroke
would gently lift them off the surface and they would continue to flap to gain
speed and elevation.
·
During
the upstroke the fore and hindwings would separate, with the forewings being
lifted up more quickly than the hindwings.
·
Flapping
frequency would probably be very low, as in larger butterflies, probably within
the range of 9-12 beats per second (Zhang et al., 2021).
·
All four
wings could be moved independently and in normal flight the flapping cycles of
the hindwings would have lagged behind the forewings.
·
During
the downstrokes the forewings would sweep down and slightly forward under the
body, and the hindwings would sweep down under the body, almost touching the
opposite wing.
·
Like
butterflies, they may have periodically glided, especially in the larger
species, but smaller species more likely employed continuous flapping like
modern neuropterans.
·
Unlike
most modern neuropterans, kalligrammatids with wings which overlap for most of
their surface probably kept their wings together on the downstroke.
·
I
imagine that they were probably quite awkward when landing, due to their large
size and relatively small legs, but this is more speculative and can’t really
be tested, It’s more of just a personal hunch.

Fig. 4 An animation showing the take-off sequence for a kalligrammatid in slow motion. Note the unfolding of the wings from over the body, the lifting up of the front legs before takeoff, and the slight delay in the flap cycle of the hindwings.
Despite
their visual similarities to butterflies, and despite often being illustrated
as almost butterfly-clones, their flight would likely have appeared quite
different. Butterflies are unusual among insects in having two pairs of wings
which function as a single unit, with the overlapping forewings providing all
of the muscle power for the downstroke; this would not have been the case for
the kalligrammatids and both fore and hind wings would have been powered during
flight. Their flight was probably more similar to other large insects, without
the erratic, floaty aspect of butterflies. However, at the end of the day, this
is all pretty speculative, and it’s difficult to predict how they would have
flown – there are no modern insects with comparably large wings except for the
butterflies and moths, so there aren’t really any good modern comparisons. The
two main takeaways from this would be that they lifter their legs up before
take-off and their wings could be moved independently, and likely did not always
function as a single unit.
Just a quick note on pollination
and mimesis
I’ve
already written far more than I intended to, and I’ve covered all of the main
points I wanted to talk about, so here’s a few quick points to wrap things up.
Firstly, pollination and feeding. As I’ve
mentioned at least twice already, most kalligrammatids had proboscises, similar
to modern butterflies and moths. These were pretty diverse and varied in both
length and how robust they were (Labandeira et al., 2016 supp. info.).
In general they were long (8 – 20 mm) and differed significantly from the proboscises
of other insects of the time (longer and more robust). Several families of Mesozoic
plants are suggested to have been hosts for the kalligrammatids; the cycads,
bennettitaleans, caytonialeans and potentially some cheirolepidiaceans (a
family of extinct conifers). Among these the bennettitaleans, specifically
members of the family Williamsoniaceae, have been suggested to be the most
likely candidates for having an association with kalligrammatids. These were shrubby
plants which produced a variety of flower-like reproductive organs which were
often large and likely produced nectars (Fig. 5). Another interesting
observation is that the proboscises of different kalligrammatids varied a lot,
with some species possessing thin and gracile proboscises, while others bore
longer, more robust proboscises; this suggest that some niche partitioning was
present in the kalligrammatids, and different species likely fed upon and
pollinated different plants (Labandeira et al., 2016).

Fig. 5 A kalligrammatid (Kalligramma brachyrhyncha) visiting a bennettitalean female cone (Wielandiella villosa) (Pott et al., 2015). Cutaway showing the internal cone structure. Note that the long robust proboscis is roughly the lame depth as the scales within the cone. Also, a cute toothy grin! Scale bar 10 mm.
Just a final quick note, before I end this far-too-long blog, is that at rest, the kalligrammatids would have folded their wings back over their bodies; this has several implications: firstly, when at rest the hindwings would have been completely or almost completely hidden by the forewings; secondly, the dorsal surfaces of the wings was the side visible when at rest (this is very different from butterflies, which fold their wings up together so that the underside of their winds is visible). This helps to explain why the eyespots of most species are present on the forewings, as they would have been hidden when at rest if on the hindwings. The undersides of the wings also may have been duller in colour, or at least may not have possessed such striking eyespots as the tops of the wings. It is also worth noting that the forewing shape in most species, and the large size of these wings roughly matches the size and shape of the heads of many small predatory dinosaurs, which are known to have inhabited the same biotas as the kalligrammatids. It seems likely then that the maniraptorans probably served as a model organism for the kalligrammatids. Furthermore, the large eyespots with round pupils and multiple rings closely matches what we predict the eyes of small maniraptorans were like (although, again, this is speculative; Fig. 6). While it seems likely that most kalligrammatids were mimicking their model organisms while at rest this does raise the question of why some species had eyespots of their hindwings. A possible, albeit speculative, explanation for this may be that species with hindwing eyespots would take flight when disturbed, suddenly exposing their previously hidden eyespots to would-be predators, startling them and scaring them off. I imagine this would be particularly affective in Makarkinia, with its massive wings and large eyespots.

That’s all folks
Well,
that’s it for now, those are all of my thoughts on the kalligrammatids, based
on the restorations I’ve recently been doing of them. Again, I just want to reiterate
that these are mostly just my own personal thoughts and observations, but I
hope that if nothing else this will help get people interested in the
kalligrammatids and restoring some more of the amazing diversity which was
present within this fascinating group.

References
Ansorge, J., Markarkin,
V. N. 2020. The oldest
giant lacewings (Neuroptera: Kalligrammatidae) from the Lower Jurassic of
Germany. Palaeoworld, vol. 30, pp. 296 – 310, doi: 10.1016/j.palwor.2020.07.001
Bechly, G., Makarkin, V.
N. 2016. A new gigantic
lacewing species (Insecta: Neuroptera) from the Lower Cretaceous of Brazil
confirms the occurrence of Kalligrammatidae in the Americas. Cretaceous Research,
vol. 58, pp. 135 – 140, doi: 10.1016/j.cretres.2015.10.014
Engel, M. S. 2005. A
remarkable kalligrammatid lacewing from the Upper Jurassic of Kazakhstan
(Neuroptera: Kalligrammatidae). Transactions of the Kansas Academy of
Science, vol. 108, iss. 1, pp. 59 – 62
Labandeira, Yang, Q., Santiago-Blay,
J. A., Hotton, C. L., Monteiro, A., Wang, Y-J., Goreva, Y., Shih, C., Siljeström,
S., Rose, T. R., Dilcher, D. L., Ren, D. 2016. The evolutionary convergence of mid-Mesozoic
lacewings and Cenozoic butterflies. Proceedings of the Royal Society B, vol.
283, art. 20152893, doi: 10.1098/rspb.2015.2893
Liu, Q., Lu, X., Zhang,
Q., Chen, J., Zheng, X., Zhang, W., Liu, X., Wang, B. 2018. High niche diversity in Mesozoic
pollinating lacewings. Nature Communications, vol. 9, art. 3793, doi: 10.1038/s41467-018-06120-5
Lu, X-M., Zhang, W-W., Liu,
X-Y. 2016. New long-proboscid
lacewings of the mid-Cretaceous provide insights into ancient plant-pollinator interactions.
Scientific Reports, vol. 6, art. 25382, doi: 10.1038/srep25382
Machado, R. J. P., Freitas,
A. V. L., Ribeiro, G. C. 2021.
A new giant species of the remarkable extinct family Kalligrammatidae (Insecta:
Neuroptera) from the Lower Cretaceous Crato Formation of Brazil. Cretaceous Research,
vol. 120, art. 104724, doi: 10.1016/j.cretres.2020.104724
Popa, M. E. 2019. Review of the bennettitalean genus Weltrichia.
Journal of Palaeogeography, vol. 8, art. 12, doi: 10.1186/s42501-019-0023-9
Pott, C., Xiaoli, W., Xiaoting,
Z. 2015. Wielandiella
villosa comb. nov. from the Middle Jurassic of Daohugou, China: More Evidence
for Divaricate Plant Architecture in Williamsoniceae. Botanica Pacifica,
vol. 4, iss. 2, pp. 137–148, doi: 10.17581/bp.2015.04115
Yang, Q., Wang, Y., Labandeira,
C. C., Shih, C., Ren, D. 2014.
Mesozoic lacewings from China provide phylogenetic insight into evolution of
the Kalligrammatidae (Neuroptera). BMC Evolutionary Biology, vol. 14,
art. 126
Zhang, Y., Wang, X., Wang,
S., Huang, W., Weng, Q. 2021.
Kinematic and Aerodynamic Investigation of the Butterfly in Forward Free Flight
for the Butterfly-Inspired Flapping Wing Air Vehicle. Applied Sciences,
vol. 11, art. 2620, doi: 10.3390/app11062620
Zhong, Y., Huyskens, M.
H., Yin, Q-Z., Wang, Y., Ma, Q., Xu, Y-G., 2021. High-precision geochronological
constraints on the duration of ‘Dinosaur Pompeii’ and the Yixian Formation. National
Science Review, vol. 8, art. nwab063, doi: 10.1093/nsr/nwab063
This is incredible work here! I'm studying to be an entomologist and I have a great interest in paleoentomology.
ReplyDeleteI am also very interested in botany. Any entomologist knows how important plants are to their field due to the incredibly large number of intimate relationships between insects and plants (mainly angiosperms).
Anyone who makes a paleontology blog that doesn't focus on vertebrates is a hero in my books! I sometimes wonder if the guys at Love in the Time of Chasmosaurs can even name five mesozoic insect or plant genera.